

概要
ゲルろ過クロマトグラフィー(以下、ゲルろ過)は、1959年にウプサラ大学の Porath と Flodin によって報告されたクロマトグラフィーの一つであり、分子の大きさの違いによってタンパク質や多糖類、核酸などの生体分子を精製する際に用いられる代表的な方法の一つである(引用1)。近年では、クライオ電子顕微鏡構造解析用のサンプルの精製や、プロテオミクス解析に向けたタンパク質複合体の精製などにおいても頻繁に用いられている。本項では、ゲルろ過の原理と実験手順、解析方法について、著者らが行ったタンパク質複合体の解析例を用いて記述した。
ゲルろ過の原理
ゲルろ過は、サンプルが多孔質の担体を通過する際に分子の大きさの違いによって溶出時間が異なることを利用した手法で、分子の大きさによってふるいにかけられたように見えることから分子ふるいとも呼ばれる(引用2)。実験上では、担体が詰まったカラムにバッファーが満たされたゲルろ過カラムとよばれる器具を用いる。原理としては、タンパク質のサイズが小さいほど、担体上の孔の内部に入り込み、担体上に滞留する時間が長くなるため、分子のサイズに反比例して溶出時間が長くなる(図1)。担体上の孔に入り込めない大きさの分子は担体上に留まることがないので、最短の時間でカラムから溶出される。このように担体によって分離できないほどの大きさのタンパク質が、カラムへロードされてから溶出されるまでの体積を排除体積(void volume)とよぶ。担体上の孔のサイズによって、分離能が異なるため目的タンパク質の大きさに対して、適切なゲルろ過カラムの選択が必要である(図2)。理論値の分子量が同じタンパク質であっても、分子の形状が異なると担体上の孔への入り込み具合が変わってくるため、溶出されるまでの時間が異なる場合がある。よくある例としては、球状タンパク質よりも細長い形状のタンパク質の方がより担体上の孔に入りにくいために、同じ分子量を有する球状タンパク質と比べて、溶出時間が短い。上記では、「溶出時間」で説明を行ったが、論文などに示すゲルろ過のデータでは、溶出体積(サンプルをロードした時点を 0 mL とし、サンプルが溶出されるまでにカラムから流れ出たバッファーの体積)を使用することが多い。また、溶出体積はカラム体積(担体の体積 + バッファーの体積)に比例することから、データの比較の際には同一のカラム体積のゲルろ過カラムを使用する必要がある。
実験背景
酵母における DNA の二重鎖切断損傷修復において、Rad51 および Rad52 は必須の因子である。二重鎖切断損傷修復の主要経路の一つである相同組換え修復機構において、Rad52 は Rad51 と相互作用し修復反応を促進すると考えられている。著者らは過去に Rad52 の C 末端領域が、Rad51 の多量体形成を阻害することを示した(引用3)。その際、Rad51 結合領域を含む Rad52 タンパク質断片(Rad52233-504)を用いて、Rad51 の会合状態の変化をゲルろ過により明らかにした。そこで本項では、著者らが過去に報告したこれらの解析をもとにゲルろ過解析の手順を解説する。
装置・器具・試薬
- Superdex200 10/300 GL(Cytiva・旧 GE Healthcare・17-5175-01)
- ÄKTA(Cytiva・旧 GE Healthcare)または NGC クロマトグラフィーシステム(Bio-Rad)
- Sample loop 500 μL(Cytiva・旧 GE Healthcare・18111399)
- 3周波超音波洗浄器 VS-100Ⅲ(AXEL・7-5345-01)
- MF-ミリポアメンブレン、0.22 μm pore size(メルクミリポア・GSWP04700)
- Gel Filtration Calibration Kit HMW(高分子用)(Cytiva・旧 GE Healthcare・28403842)
ランニングバッファー
- 20 mM HEPES-KOH(pH 7.5)
- 0.1 M KCl
- 0.5 mM ethylenediaminetetraacetic acid(EDTA)
- 10% glycerol
- 2 mM 2-mercaptoethanol
タンパク質溶液の組成
- Rad52233-504
- 20 mM HEPES-KOH(pH 7.5)
- 0.15 M KCl
- 0.5 mM EDTA
- 10% glycerol
- 2 mM 2-mercaptoethanol
- Rad51
- 20 mM Tris-HCl(pH 7.5)
- 420 mM KCl
- 0.2 mM EDTA
- 5 mM 2-mercaptoethanol
- 5% glycerol
- 0.2 mM PMSF
タンパク質の調製
本実験では、終濃度 20 μM となるように希釈した Rad52233-504 ならびに終濃度 10 μM となるように希釈した Rad51 を用いてゲルろ過の解析を行なった。Rad52233-504–Rad51 複合体については、Rad52233-504(終濃度 20 μM)と Rad51(終濃度 10 μM)を直接混合し、室温で1時間置いたものをゲルろ過解析に用いた。詳細は Kagawa et al., Nucleic Acids Research, 2014(引用3)を参照。
主な実験手順
- カラムの平衡化
- サンプルのロード
- サンプルの溶出
- カラムの洗浄
- サンプルのゲルろ過解析
実験の詳細
1. カラムの平衡化
サンプルをロードする前にはランニングバッファーをゲルろ過カラムに流し、ゲルろ過カラムの平衡化を行う必要がある。通常、ゲルろ過カラムの平衡化にはカラム体積の1.5倍以上のバッファーを使用する。クロマトグラフィーシステムの流路に沈殿物や気泡を入れないように、バッファーは使用する前に必ずフィルターを通し脱気する必要がある。沈殿物は流路やカラム上部のフィルターに詰まってしまい、カラムにかかる圧力の上昇の原因になる。また、気泡は流路やカラム内に留まることで圧力が不安定になり、カラムの劣化や結果の再現性の低下につながる。一般的に、バッファーは 0.22 μm メンブレンのフィルターにかけ、脱気は15~30分程度行えば十分である。著者らは、脱気を促すために、バッファーの入った容器に超音波を当てながら減圧を行なっている。フィルターにかけた後は、バッファーにほこりなどが混入しないよう注意して取り扱う必要がある。また、バッファーとクロマトグラフィーシステムの間に温度差がある状態でバッファーをシステムの中に流すと、システムやカラム内に気泡が発生する原因となることから、バッファーの温度をシステム内の温度と同じにしてから使用する。通常は、タンパク質の安定性を考慮し、4℃下でゲルろ過を行う。
バッファーをゲルろ過カラムに流す際は、必ずカラムにかかる圧力を確認しながら行う。各カラムのマニュアルには限界圧(pressure limit)と推奨の流速が記載されているが、カラムにかかる実際の圧力はバッファーの粘性や、カラムの汚れ具合によって大きく異なる。また、カラム内部の溶液を、組成が大きく異なる溶液に置換する場合は、カラム内の圧力が急上昇することがあるので、定期的に圧力を確認する必要がある。本項で用いた ÄKTA や Bio-Rad 社のクロマトグラフィーシステム(NGC Medium-Pressure Chromatography System など)では、圧力の上限及び下限を設定することができる。カラムの破損などを防ぐためにも、この機能を使用することを推奨する。また、再現性の高いゲルろ過解析を行うためには、クロマトグラフィーを止めることなくサンプルの溶出を完遂することが重要である。そのため著者らは、平衡化中のカラム圧力値を参考にして流速を調整し、限界圧の60%~70%程度になる流速で以下の操作を行なっている。
2. サンプルのロード
ゲルろ過カラムにロードできるサンプルの体積は、カラム体積に依存する。ゲルろ過カラムの分離能は、ロードするサンプルの体積が大きすぎると低下する。Cytiva(旧 GE Healthcare)はカラム体積の4%以下の体積のサンプル量であれば分離が可能としており、より良い分離能を得るためには、サンプル量をカラム体積の2%以下にすることを推奨している。なお、著者らがゲルろ過を行う際には、通常サンプル量をカラム体積の1%程度としている。サンプルはフィルター(5~0.22 μm)を通すか、遠心(2,000‒3,000 × g, 5 min, 4℃)してその上清を回収することにより、不溶性の沈殿物を取り除く。本項の実験では、Superdex 200 10/300 GL ゲルろ過カラム(カラム体積が約 24 mL)を使用し、検量線用の分子量マーカーも含めて全てのサンプルの体積を 250 μL になるように調製した。本実験では 500 μL サンプルループを使用し、サンプル溶出時の最初の 1 mL はサンプルループを経由するように設定することで、サンプルループや流路にサンプルが残らないようにした。
3. サンプルの溶出
ÄKTA や NGC クロマトグラフィーシステムを用いてゲルろ過カラムから溶出されたサンプルを分画する際に、フラクションの体積に注意する必要がある。ÄKTA や NGC クロマトグラフィーシステムでは、溶出液の吸光度(280 nm および 260 nm)を連続的に測定するので、通常、その情報を元にタンパク質が含まれていると予想されるフラクションの一部を SDS-PAGE で分離し、目的タンパク質が含まれているかを調べる。本項ではフラクションの体積を 500 μL に設定した。1つの溶出ピーク内においても、溶出体積の違いによってタンパク質の純度・形状・活性などが異なる場合がある。そのため、目的に応じてフラクションの体積を変える必要がある。例えば、なるべく濃度が高いサンプルが必要な場合は、各フラクションの体積を小さく(例:150 μL)設定することで溶出ピークの頂点付近のフラクションを回収することができる。また、クロマトグラフィーシステムで回収できるフラクションの数には限りがあることから、カラム体積の大きいカラム(HiLoad など)を用いる際には、排除体積からフラクションの採取を開始したり、溶出ピークのみを採取するように設定することで、目的サンプルの溶出ピークを細かく分画することができる。
4. カラムの洗浄
Cytiva 社のマニュアルでは、Superdex シリーズのゲルろ過カラムを10~20回使用する毎に NaOH 溶液で洗浄することを推奨している。今回の実験では、異なるサンプルの混入の危険性を防ぐために、各サンプルをロードする前にカラムの洗浄を行った。そのため、同じタンパク質を複数回に分けて精製する際には、以下のカラムの洗浄を必ずしも行う必要はない。通常の Superdex の洗浄(マニュアルに記載)では、まず1カラム体積分以上の Milli-Q 水をカラムへ流し、バッファーをカラムから洗い流す。次に1カラム体積分の 0.5 M NaOH を流したのちに、2カラム体積以上の Milli-Q 水をカラムに流す。一方、筆者らはカラムを簡易的に洗浄するため、カラムへ 0.5 M NaOH を 5 mL 程度ロードした後に、1カラム体積分の Milli-Q 水で洗浄した。その後、2カラム体積分のランニングバッファーでカラムを再平衡化し、次のサンプルの解析を行った。なお、洗浄に用いる溶液や手法については、各カラム担体によって多少異なるので、必ずマニュアルを確認することを勧める。
5. サンプルのゲルろ過解析
上記の方法で、Rad52 C 末端側断片(Rad52233-504)、Rad51、Rad52233-504 と Rad51 混合溶液のゲルろ過解析を行った結果を示す(図3A)。Rad52233-504 は 13 mL 付近に、Rad51 は 11 mL 付近に溶出されたが、Rad52233-504 と Rad51 を混合した際には、12 mL 付近に主要な溶出ピークが検出された。混合液にて検出された溶出ピークは、Rad52233-504 単体よりも溶出体積が小さかったことから、Rad52233-504 は Rad51 と相互作用したことが考えられた。実際、混合溶液の溶出ピークを SDS-PAGE で解析した結果、Rad52233-504 と Rad51 両方のタンパク質が検出され(図3B)、Rad52233-504 と Rad51 が複合体を形成していることが示唆された。また一方で、Rad51 単独時の溶出ピークは Rad51 単量体の分子量から推測される溶出体積よりもはるかに小さい体積で溶出しており、これまでの報告と同様に Rad51 が多量体を形成していることがわかる。しかし、混合液にて検出された溶出ピークはこの Rad51 多量体の溶出ピークより溶出体積が大きく、Rad52233-504 が Rad51 と結合すると、Rad51 どうしの会合状態が変化もしくは会合そのものが阻害されることが示唆された(引用3)。
ゲルろ過解析により分子量を求めるには、分子量が既知のタンパク質(分子量マーカー)の溶出体積から作成した検量線を用いる。ÄKTA クロマトグラフィーシステムの場合、UNICORN software 内のpeak integrate の機能を使用すると容易に溶出体積を求めることができる。排除体積の検出には、分子量マーカーのキットに含まれている Blue Dextran を使用した。縦軸は分子量(対数目盛)、横軸を以下の式から算出した \(\mathrm{K_{av}}\) 値をプロットし、その近似直線を検量線とした(図4)。検量線の式に目的タンパク質の \(\mathrm{K_{av}}\) 値を代入することにより、おおよその分子量を算出することができる。なお、ランニングバッファーの組成および流速は溶出体積に影響を与える可能性があるため、分子量マーカーの解析は目的サンプルと同じ条件で行う必要がある。
\[ \mathrm{K_{av}} = \frac{\text{(溶出体積)} − \text{(排除体積)}}{\text{(カラム体積)} − \text{(排除体積)}} \]
ゲルろ過解析により求めたRad52233-504、Rad51、Rad52233-504–Rad51 複合体の分子量を示す(表1)。さらにゲルろ過によって精製したサンプルは超遠心分析でも分子量の算出を行なった。なお、Rad51 は会合状態が不均一であるため、超遠心分析を行っていない。ゲルろ過で求めた分子量は、アミノ酸配列から算出されたタンパク質の理論値に比べて、かなり大きかった。このような場合には、タンパク質が会合状態を形成していることや球状タンパク質とは異なる形状の構造を有していることが考えられる。しかし、超遠心分析の結果では、Rad52233-504 および Rad52233-504–Rad51 複合体は理論値に近い値であることから、溶液中において、Rad52233-504 は単量体、Rad52233-504–Rad51 複合体はヘテロ二量体を形成し、その構造は球状タンパク質とは大きく異なることが考えられた。
このようにゲルろ過は、溶出体積や溶出ピークの形状を調べることで、比較的容易に標的タンパク質の構造の均一性、分子量、複合体形成を解析することができる。近年、クライオ電子顕微鏡によるタンパク質の構造解析技術の著しい発展や、プロテオミクス解析の感度向上に伴い、精製度の高いタンパク質(もしくはタンパク質複合体)の調製が求められる機会が増えている。そうした状況の中、比較的古い実験手法であるゲルろ過の有用性が再認識されている。
表1:各解析法により求められるタンパク質の分子量
| ゲルろ過解析(kDa) | 超遠心分析(kDa) | 理論値(kDa) | |
|---|---|---|---|
| Rad52233-504 | 122.7 | 28.9 | 30.1 |
| Rad51 | 298.6 | –‒ | 42.9 |
| Rad52233-504–Rad51 | 181.5 | 73.1 | 73.1 |
タンパク質の理論値については、各タンパク質のアミノ酸配列をもとにして ExPASy の ProtParam tool を使用して求めた(https://web.expasy.org/protparam/)。
文献
- 岡澤 敦司, 生物工学会誌, 93(6):345–48, ザ・ヒストリー・オブ・クロマトグラフィー (2015)
- Cytiva(旧 GE Healthcare), Gel filtration principles and Method (2003)
- Kagawa W., Arai N., Ichikawa Y., Saito K., Sugiyama S., Saotome M., Shibata T. and Kurumizaka H., Nucleic Acids Research, 42(2):941‒951 (2014)
- ゲル濾過担体選択ガイド(Cytiva), https://www.cytivalifesciences.co.jp/newsletter/biodirect_mail/technical_tips/pdf/selection_guide.pdf
-

図1:ゲルろ過解析の原理。(A)ゲルろ過カラムの模式図。灰色は担体。青色はバッファーを示している。カラム体積とは、担体の体積とバッファーの体積を合わせた体積のことを指す。なお、排除体積は、バッファーの体積に相当する。担体表面は多数の小さな穴が空いた多孔質の素材からできている。(B)ゲルろ過における分子ふるいの原理に関する模式図。タンパク質がカラムを通る際、担体上に自身が入れる大きさの孔が存在すると入り込こむ。そのため、タンパク質が小さいほど孔の中での滞在時間が長くなることから、溶出体積は大きくなる。(C)B においてそれぞれのタンパク質が通る経路を示した図。分子の小さいタンパク質ほど流路が長くなる。 -
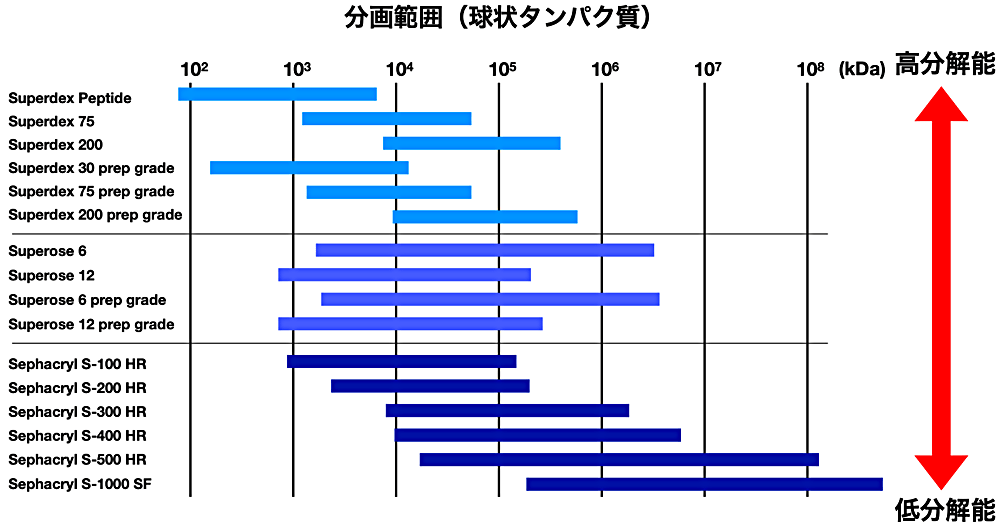
図2:ゲルろ過カラムの分離能。ゲルろ過解析はタンパク質の構造に依存して、理論的な分子量から予測される溶出体積とは異なる体積で溶出される場合もある。よって、ゲルろ過カラムを選択する際は、目的タンパク質の分子量の理論値が、ゲルろ過カラムの分画範囲の中間あたりになるようなカラムを選択すると良い。ゲルろ過担体選択ガイド(文献4)を参照。 -

図3:Rad52233-504、Rad51 および複合体のゲルろ過解析。(A)Rad52233-504、Rad51 および Rad52233-504–Rad51 複合体のクロマトグラム。分子量の目盛りは、分子量マーカーの溶出体積を示す。(B)(A)で四角で囲われた範囲のフラクションの電気泳動図。混合液の溶出ピークには Rad52233-504 と Rad51 が含まれていることがわかった。Kagawa et al., Nucleic Acids Research (2013)(引用3)より引用、一部改変。 -
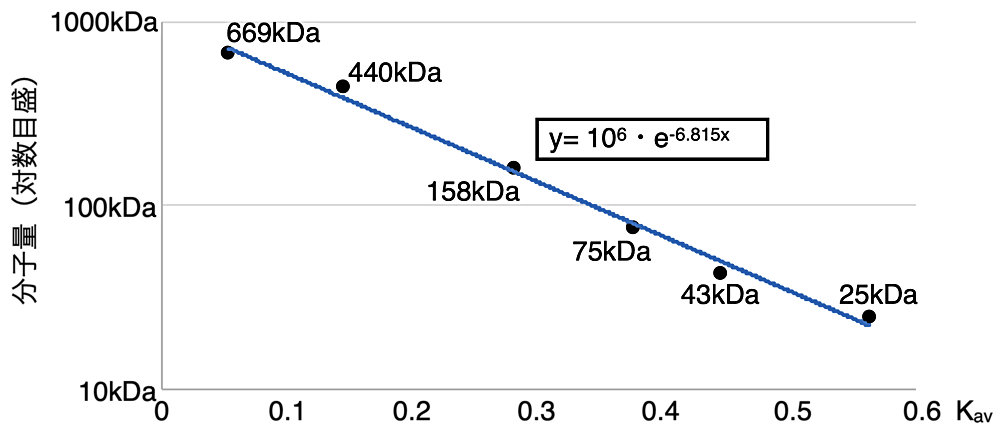
図4:分子量マーカーから求めた検量線。分子量マーカーの溶出体積をもとに作成した検量線(Microsoft Excel を使用)。得られた近似直線の式に、分子量を求めたいタンパク質の \(\mathrm{K_{av}}\) を代入することで、分子量を求めることができる。
概要
ゲルろ過クロマトグラフィー(以下、ゲルろ過)は、1959年にウプサラ大学の Porath と Flodin によって報告されたクロマトグラフィーの一つであり、分子の大きさの違いによってタンパク質や多糖類、核酸などの生体分子を精製する際に用いられる代表的な方法の一つである(引用1)。近年では、クライオ電子顕微鏡構造解析用のサンプルの精製や、プロテオミクス解析に向けたタンパク質複合体の精製などにおいても頻繁に用いられている。本項では、ゲルろ過の原理と実験手順、解析方法について、著者らが行ったタンパク質複合体の解析例を用いて記述した。
ゲルろ過の原理
ゲルろ過は、サンプルが多孔質の担体を通過する際に分子の大きさの違いによって溶出時間が異なることを利用した手法で、分子の大きさによってふるいにかけられたように見えることから分子ふるいとも呼ばれる(引用2)。実験上では、担体が詰まったカラムにバッファーが満たされたゲルろ過カラムとよばれる器具を用いる。原理としては、タンパク質のサイズが小さいほど、担体上の孔の内部に入り込み、担体上に滞留する時間が長くなるため、分子のサイズに反比例して溶出時間が長くなる(図1)。担体上の孔に入り込めない大きさの分子は担体上に留まることがないので、最短の時間でカラムから溶出される。このように担体によって分離できないほどの大きさのタンパク質が、カラムへロードされてから溶出されるまでの体積を排除体積(void volume)とよぶ。担体上の孔のサイズによって、分離能が異なるため目的タンパク質の大きさに対して、適切なゲルろ過カラムの選択が必要である(図2)。理論値の分子量が同じタンパク質であっても、分子の形状が異なると担体上の孔への入り込み具合が変わってくるため、溶出されるまでの時間が異なる場合がある。よくある例としては、球状タンパク質よりも細長い形状のタンパク質の方がより担体上の孔に入りにくいために、同じ分子量を有する球状タンパク質と比べて、溶出時間が短い。上記では、「溶出時間」で説明を行ったが、論文などに示すゲルろ過のデータでは、溶出体積(サンプルをロードした時点を 0 mL とし、サンプルが溶出されるまでにカラムから流れ出たバッファーの体積)を使用することが多い。また、溶出体積はカラム体積(担体の体積 + バッファーの体積)に比例することから、データの比較の際には同一のカラム体積のゲルろ過カラムを使用する必要がある。
実験背景
酵母における DNA の二重鎖切断損傷修復において、Rad51 および Rad52 は必須の因子である。二重鎖切断損傷修復の主要経路の一つである相同組換え修復機構において、Rad52 は Rad51 と相互作用し修復反応を促進すると考えられている。著者らは過去に Rad52 の C 末端領域が、Rad51 の多量体形成を阻害することを示した(引用3)。その際、Rad51 結合領域を含む Rad52 タンパク質断片(Rad52233-504)を用いて、Rad51 の会合状態の変化をゲルろ過により明らかにした。そこで本項では、著者らが過去に報告したこれらの解析をもとにゲルろ過解析の手順を解説する。
装置・器具・試薬
- Superdex200 10/300 GL(Cytiva・旧 GE Healthcare・17-5175-01)
- ÄKTA(Cytiva・旧 GE Healthcare)または NGC クロマトグラフィーシステム(Bio-Rad)
- Sample loop 500 μL(Cytiva・旧 GE Healthcare・18111399)
- 3周波超音波洗浄器 VS-100Ⅲ(AXEL・7-5345-01)
- MF-ミリポアメンブレン、0.22 μm pore size(メルクミリポア・GSWP04700)
- Gel Filtration Calibration Kit HMW(高分子用)(Cytiva・旧 GE Healthcare・28403842)
ランニングバッファー
- 20 mM HEPES-KOH(pH 7.5)
- 0.1 M KCl
- 0.5 mM ethylenediaminetetraacetic acid(EDTA)
- 10% glycerol
- 2 mM 2-mercaptoethanol
タンパク質溶液の組成
- Rad52233-504
- 20 mM HEPES-KOH(pH 7.5)
- 0.15 M KCl
- 0.5 mM EDTA
- 10% glycerol
- 2 mM 2-mercaptoethanol
- Rad51
- 20 mM Tris-HCl(pH 7.5)
- 420 mM KCl
- 0.2 mM EDTA
- 5 mM 2-mercaptoethanol
- 5% glycerol
- 0.2 mM PMSF
タンパク質の調製
本実験では、終濃度 20 μM となるように希釈した Rad52233-504 ならびに終濃度 10 μM となるように希釈した Rad51 を用いてゲルろ過の解析を行なった。Rad52233-504–Rad51 複合体については、Rad52233-504(終濃度 20 μM)と Rad51(終濃度 10 μM)を直接混合し、室温で1時間置いたものをゲルろ過解析に用いた。詳細は Kagawa et al., Nucleic Acids Research, 2014(引用3)を参照。
主な実験手順
- カラムの平衡化
- サンプルのロード
- サンプルの溶出
- カラムの洗浄
- サンプルのゲルろ過解析
実験の詳細
1. カラムの平衡化
サンプルをロードする前にはランニングバッファーをゲルろ過カラムに流し、ゲルろ過カラムの平衡化を行う必要がある。通常、ゲルろ過カラムの平衡化にはカラム体積の1.5倍以上のバッファーを使用する。クロマトグラフィーシステムの流路に沈殿物や気泡を入れないように、バッファーは使用する前に必ずフィルターを通し脱気する必要がある。沈殿物は流路やカラム上部のフィルターに詰まってしまい、カラムにかかる圧力の上昇の原因になる。また、気泡は流路やカラム内に留まることで圧力が不安定になり、カラムの劣化や結果の再現性の低下につながる。一般的に、バッファーは 0.22 μm メンブレンのフィルターにかけ、脱気は15~30分程度行えば十分である。著者らは、脱気を促すために、バッファーの入った容器に超音波を当てながら減圧を行なっている。フィルターにかけた後は、バッファーにほこりなどが混入しないよう注意して取り扱う必要がある。また、バッファーとクロマトグラフィーシステムの間に温度差がある状態でバッファーをシステムの中に流すと、システムやカラム内に気泡が発生する原因となることから、バッファーの温度をシステム内の温度と同じにしてから使用する。通常は、タンパク質の安定性を考慮し、4℃下でゲルろ過を行う。
バッファーをゲルろ過カラムに流す際は、必ずカラムにかかる圧力を確認しながら行う。各カラムのマニュアルには限界圧(pressure limit)と推奨の流速が記載されているが、カラムにかかる実際の圧力はバッファーの粘性や、カラムの汚れ具合によって大きく異なる。また、カラム内部の溶液を、組成が大きく異なる溶液に置換する場合は、カラム内の圧力が急上昇することがあるので、定期的に圧力を確認する必要がある。本項で用いた ÄKTA や Bio-Rad 社のクロマトグラフィーシステム(NGC Medium-Pressure Chromatography System など)では、圧力の上限及び下限を設定することができる。カラムの破損などを防ぐためにも、この機能を使用することを推奨する。また、再現性の高いゲルろ過解析を行うためには、クロマトグラフィーを止めることなくサンプルの溶出を完遂することが重要である。そのため著者らは、平衡化中のカラム圧力値を参考にして流速を調整し、限界圧の60%~70%程度になる流速で以下の操作を行なっている。
2. サンプルのロード
ゲルろ過カラムにロードできるサンプルの体積は、カラム体積に依存する。ゲルろ過カラムの分離能は、ロードするサンプルの体積が大きすぎると低下する。Cytiva(旧 GE Healthcare)はカラム体積の4%以下の体積のサンプル量であれば分離が可能としており、より良い分離能を得るためには、サンプル量をカラム体積の2%以下にすることを推奨している。なお、著者らがゲルろ過を行う際には、通常サンプル量をカラム体積の1%程度としている。サンプルはフィルター(5~0.22 μm)を通すか、遠心(2,000‒3,000 × g, 5 min, 4℃)してその上清を回収することにより、不溶性の沈殿物を取り除く。本項の実験では、Superdex 200 10/300 GL ゲルろ過カラム(カラム体積が約 24 mL)を使用し、検量線用の分子量マーカーも含めて全てのサンプルの体積を 250 μL になるように調製した。本実験では 500 μL サンプルループを使用し、サンプル溶出時の最初の 1 mL はサンプルループを経由するように設定することで、サンプルループや流路にサンプルが残らないようにした。
3. サンプルの溶出
ÄKTA や NGC クロマトグラフィーシステムを用いてゲルろ過カラムから溶出されたサンプルを分画する際に、フラクションの体積に注意する必要がある。ÄKTA や NGC クロマトグラフィーシステムでは、溶出液の吸光度(280 nm および 260 nm)を連続的に測定するので、通常、その情報を元にタンパク質が含まれていると予想されるフラクションの一部を SDS-PAGE で分離し、目的タンパク質が含まれているかを調べる。本項ではフラクションの体積を 500 μL に設定した。1つの溶出ピーク内においても、溶出体積の違いによってタンパク質の純度・形状・活性などが異なる場合がある。そのため、目的に応じてフラクションの体積を変える必要がある。例えば、なるべく濃度が高いサンプルが必要な場合は、各フラクションの体積を小さく(例:150 μL)設定することで溶出ピークの頂点付近のフラクションを回収することができる。また、クロマトグラフィーシステムで回収できるフラクションの数には限りがあることから、カラム体積の大きいカラム(HiLoad など)を用いる際には、排除体積からフラクションの採取を開始したり、溶出ピークのみを採取するように設定することで、目的サンプルの溶出ピークを細かく分画することができる。
4. カラムの洗浄
Cytiva 社のマニュアルでは、Superdex シリーズのゲルろ過カラムを10~20回使用する毎に NaOH 溶液で洗浄することを推奨している。今回の実験では、異なるサンプルの混入の危険性を防ぐために、各サンプルをロードする前にカラムの洗浄を行った。そのため、同じタンパク質を複数回に分けて精製する際には、以下のカラムの洗浄を必ずしも行う必要はない。通常の Superdex の洗浄(マニュアルに記載)では、まず1カラム体積分以上の Milli-Q 水をカラムへ流し、バッファーをカラムから洗い流す。次に1カラム体積分の 0.5 M NaOH を流したのちに、2カラム体積以上の Milli-Q 水をカラムに流す。一方、筆者らはカラムを簡易的に洗浄するため、カラムへ 0.5 M NaOH を 5 mL 程度ロードした後に、1カラム体積分の Milli-Q 水で洗浄した。その後、2カラム体積分のランニングバッファーでカラムを再平衡化し、次のサンプルの解析を行った。なお、洗浄に用いる溶液や手法については、各カラム担体によって多少異なるので、必ずマニュアルを確認することを勧める。
5. サンプルのゲルろ過解析
上記の方法で、Rad52 C 末端側断片(Rad52233-504)、Rad51、Rad52233-504 と Rad51 混合溶液のゲルろ過解析を行った結果を示す(図3A)。Rad52233-504 は 13 mL 付近に、Rad51 は 11 mL 付近に溶出されたが、Rad52233-504 と Rad51 を混合した際には、12 mL 付近に主要な溶出ピークが検出された。混合液にて検出された溶出ピークは、Rad52233-504 単体よりも溶出体積が小さかったことから、Rad52233-504 は Rad51 と相互作用したことが考えられた。実際、混合溶液の溶出ピークを SDS-PAGE で解析した結果、Rad52233-504 と Rad51 両方のタンパク質が検出され(図3B)、Rad52233-504 と Rad51 が複合体を形成していることが示唆された。また一方で、Rad51 単独時の溶出ピークは Rad51 単量体の分子量から推測される溶出体積よりもはるかに小さい体積で溶出しており、これまでの報告と同様に Rad51 が多量体を形成していることがわかる。しかし、混合液にて検出された溶出ピークはこの Rad51 多量体の溶出ピークより溶出体積が大きく、Rad52233-504 が Rad51 と結合すると、Rad51 どうしの会合状態が変化もしくは会合そのものが阻害されることが示唆された(引用3)。
ゲルろ過解析により分子量を求めるには、分子量が既知のタンパク質(分子量マーカー)の溶出体積から作成した検量線を用いる。ÄKTA クロマトグラフィーシステムの場合、UNICORN software 内のpeak integrate の機能を使用すると容易に溶出体積を求めることができる。排除体積の検出には、分子量マーカーのキットに含まれている Blue Dextran を使用した。縦軸は分子量(対数目盛)、横軸を以下の式から算出した \(\mathrm{K_{av}}\) 値をプロットし、その近似直線を検量線とした(図4)。検量線の式に目的タンパク質の \(\mathrm{K_{av}}\) 値を代入することにより、おおよその分子量を算出することができる。なお、ランニングバッファーの組成および流速は溶出体積に影響を与える可能性があるため、分子量マーカーの解析は目的サンプルと同じ条件で行う必要がある。
\[ \mathrm{K_{av}} = \frac{\text{(溶出体積)} − \text{(排除体積)}}{\text{(カラム体積)} − \text{(排除体積)}} \]
ゲルろ過解析により求めたRad52233-504、Rad51、Rad52233-504–Rad51 複合体の分子量を示す(表1)。さらにゲルろ過によって精製したサンプルは超遠心分析でも分子量の算出を行なった。なお、Rad51 は会合状態が不均一であるため、超遠心分析を行っていない。ゲルろ過で求めた分子量は、アミノ酸配列から算出されたタンパク質の理論値に比べて、かなり大きかった。このような場合には、タンパク質が会合状態を形成していることや球状タンパク質とは異なる形状の構造を有していることが考えられる。しかし、超遠心分析の結果では、Rad52233-504 および Rad52233-504–Rad51 複合体は理論値に近い値であることから、溶液中において、Rad52233-504 は単量体、Rad52233-504–Rad51 複合体はヘテロ二量体を形成し、その構造は球状タンパク質とは大きく異なることが考えられた。
このようにゲルろ過は、溶出体積や溶出ピークの形状を調べることで、比較的容易に標的タンパク質の構造の均一性、分子量、複合体形成を解析することができる。近年、クライオ電子顕微鏡によるタンパク質の構造解析技術の著しい発展や、プロテオミクス解析の感度向上に伴い、精製度の高いタンパク質(もしくはタンパク質複合体)の調製が求められる機会が増えている。そうした状況の中、比較的古い実験手法であるゲルろ過の有用性が再認識されている。
表1:各解析法により求められるタンパク質の分子量
| ゲルろ過解析(kDa) | 超遠心分析(kDa) | 理論値(kDa) | |
|---|---|---|---|
| Rad52233-504 | 122.7 | 28.9 | 30.1 |
| Rad51 | 298.6 | –‒ | 42.9 |
| Rad52233-504–Rad51 | 181.5 | 73.1 | 73.1 |
タンパク質の理論値については、各タンパク質のアミノ酸配列をもとにして ExPASy の ProtParam tool を使用して求めた(https://web.expasy.org/protparam/)。
文献
- 岡澤 敦司, 生物工学会誌, 93(6):345–48, ザ・ヒストリー・オブ・クロマトグラフィー (2015)
- Cytiva(旧 GE Healthcare), Gel filtration principles and Method (2003)
- Kagawa W., Arai N., Ichikawa Y., Saito K., Sugiyama S., Saotome M., Shibata T. and Kurumizaka H., Nucleic Acids Research, 42(2):941‒951 (2014)
- ゲル濾過担体選択ガイド(Cytiva), https://www.cytivalifesciences.co.jp/newsletter/biodirect_mail/technical_tips/pdf/selection_guide.pdf
